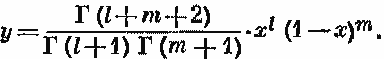Размещение растений в пределах сообществ. Использование отрицательного биномиального распределения.
Использование отрицательного биномиального распределения также основано на недоказанных допущениях, касающихся природы скоплений особей. Это распределение характеризуется средним числом особей на площадку и показателем степени, зависящим от средней скорости возобновления. Здесь снова вызывает сомнение возможность использования оценки второго параметра. Робинсон, исходя из теоретических предпосылок, утверждает, что величина показателя должна быть независимой от внешних условий и, следовательно, для одного и того же вида одинаковой в разных сообществах. Материал, подкрепляющий это утверждение, он находит в данных Стейгера. Однако не ясно, почему характер возобновления должен оставаться одинаковым в разных условиях; в действительности все то, что известно о растениях в природе, позволяет предполагать, что он меняется. В этом отношении данные Стейгера имеют небольшое значение по причинам, отмеченным выше. Далее, положение о неизменности значения показателя для любого отдельного вида не подтверждается данными Пиджеона и Эшби, показавшими, что размещение вида может быть случайным на одном участке и явно групповым — на другом.
При сравнении предложенных разными авторами распределений следует помнить, что существует очень много теоретически возможных распределений этого рода.
Поэтому удовлетворительное соответствие полученных данных одному из них едва ли можно рассматривать как доказательство его преимущественной пригодности, если отсутствуют веские и полученные независимым путем биологические свидетельства справедливости допущений, лежащих в его основе.
Важность различий условий среды в пределах исследуемого участка подчеркивается тем фактом, что удовлетворительное соответствие распределениям Неймана и Томаса было получено только в тех случаях, когда эти различия были сведены к минимуму. Арчибальд использовал для получения своих данных 100 или 500 смежных площадок, каждая площадью 20 см2, так что даже при наблюдениях с 500-кратной повторностью общая площадь, с которой отбирались образцы, составляла всего 1 м2. Такая площадь столь мала, что изменчивость условий среды на ней не проявляется, особенно в таких относительно однородных местообитаниях, как изучавшиеся автором засоленные марши и поросшие травой территории, подстилаемые мелом. Варне и Стэнбери намеренно избрали объектом работы исключительно однородное местообитание (площадки, образующиеся при осаждении за искусственными дамбами каолина, вынесенного из карьеров) и изучали только ранние стадии сукцессии, происходящей благодаря заносу зачатков водой и ветром.
Единственный опыт математической характеристики неслучайного размещения на основе первого механизма, т. е. изменчивости пуассоновского параметра в пределах участка, или изменчивости условий среды, был, по-видимому. предпринят Стивенсом. Он вывел формулу
![]()
где Е — число «пустых» квадратов, ожидаемое на учетной площадке, разделенной на п квадратов меньшей площади, причем на площадке насчитывается s особей, распределенных случайно.
Если распределение не является случайным, он предложил использовать видоизмененную формулу:
![]()
где с рассчитывается таким образом, чтобы получить наилучшее соответствие данным наблюдений. Следовательно, с является мерой изменчивости пуассоновского параметра в пределах участка. Этот метод был применен Эшби при исследовании группировки Salicornia europaea на солончаковых маршах и Сингхом и Дасом при изучении распространения сорняка Bonnaya brachiata на пахотных землях в Индии. Ни в том, ни в другом случае не было проведено какой-либо проверки степени согласия, получаемой с помощью уточненной формулы для Е, однако в обоих случаях она объясняет значительную часть отклонения от случайности. Хотя этот подход основан на учете изменчивости, связанной с колебаниями условий среды, в обоих случаях, где он был применен, использовались пробные площадки таких размеров, что с их помощью можно было обнаружить лишь гетерогенность низшего порядка, возникающую благодаря существованию мелких групп особей. Такие группы образуются в процессе размножения растений. Другими словами, хотя ставилась задача исследовать первый механизм (влияние изменчивости среды), фактически исследовался второй (влияние характера возобновления). Таким образом, вновь подтверждается мнение Феллера, считающего, что разделить действие двух механизмов невозможно. Теоретически вполне можно было бы сделать пробную площадку значительно больше, что позволило бы уловить изменчивость местообитания. Однако для всех видов, кроме видов с очень низкой численностью, для получения достаточного числа «пустых» квадратов потребуется заложить такое большое число квадратов внутри площадки, что их учет займет очень много времени и метод станет практически невыгодным. Другое ограничение состоит в том, что метод применим лишь в тех случаях,
когда с мало по отношению к средней вероятности встречи особи. Наконец, показатель с трудно использовать для характеристики свойств как растения, так и местообитания.
Эриксон и Стэн выдвинули предложение, согласно которому местообитание можно разделить на две части: благоприятную для изучаемого вида и неблагоприятную или по крайней мере не занятую видом.
Тогда результаты наблюдений можно соответственно разделить на две группы, одна из которых будет характеризовать благоприятную, другая — неблагоприятную часть местообитания. Чтобы произвести такое деление, Эриксон и Стэн построили график зависимости log (x!y) от х (у — число пробных площадок, содержащих х особей). Вычерченный таким образом график пуассоновского распределения имеет вид прямой:
log (xly) = log (Ne-m) + x log m
так как для N площадок со средним, равным.
Авторы на глаз довели прямую до точек, соответствующих наиболее высоким значениям х, и по полученному графику определили число площадок, которые должны быть отнесены к «благоприятной» части участка, а также среднюю численность на этих площадках. Распределение остальных площадок, которые относили к «неблагоприятной» части, в большинстве случаев обнаружило хорошее соответствие с пуассоновским распределением. Этот подход, на который не было обращено должного внимания исследователей, ценен тем, что концентрирует внимание на изменчивости пуассоновского параметра, но применимость его ограниченна, так как он основан на допущении, что пуассоновский параметр характеризуется прерывистой изменчивостью, тогда как во многих случаях это маловероятно. При наличии пятнистости, вызываемой особенностями размножения некоторых видов растений, этот подход непригоден.
Кемп и Кемп рассматривали вопрос о распределении, ожидаемом при учете покрытия с помощью рамок с иглами.
Исходя из предположения Робинсона, согласно которому для покрытия следует ожидать бета распределения [1] они построили распределение, приняв, что вероятность укола иглой для отдельного вида в пределах одной рамки постоянна, но в пределах совокупности рамок подчиняется бета-распределению.
Получаемое в результате распределение можно описать с помощью среднего и дисперсии бета-распределения; последняя служит мерой вариабельности показаний отдельных рамок, т. е. связана с различиями условий среды в разных точках участка. Авторы нашли, что полученное ими распределение удовлетворительно согласуется с данными по покрытию, взятыми у Гудолл а. Однако и здесь лежащие в основе теоретического построения допущения могут быть оспариваемы. Прежде всего не было представлено реальных доказательств приемлемости бета-распределения. Далее, сомнительно, чтобы для обычных типов рамок вероятность внутри рамки оставалась постоянной, хотя это возражение можно устранить, употребляя рамку меньшего размера. Наконец, как и в случае с методом Стивенса, второй параметр, хотя он и характеризует изменчивость условий в отдельных точках участка, трудно интерпретировать с биологической точки зрения.
Подводя итоги рассмотренных попыток использовать модификации пуассоновского или биномиального распределений для изучения отклонений от случайности, обнаруживаемых в полевых данных, можно прийти к выводу, что они приводят либо к подтверждению очевидного, т. е. что вид имеет тенденцию к произрастанию компактными группами из нескольких особей, либо к определению типа размещения с помощью констант, которым нельзя приписать какое-либо точное биологическое значение. По-видимому, более эмпирический подход, хотя он и не удовлетворит математически мыслящего исследователя, принесет больше пользы геоботаникам, которым всегда, кроме самых благоприятных случаев, приходится сталкиваться с большим числом переменных, требующих учета.
Различные исследования типов размещения, произведенные до сих пор, дают возможность сделать два вывода:
- во-первых, виды относительно редко обнаруживают случайное распределение даже в пределах мелких однородных участков;
- во-вторых, групповое (кучное) распределение особей встречается гораздо чаще, чем равномерное.
Это кажется на первый взгляд неожиданным, так как можно было бы ожидать, что следствием конкуренции между особями должно быть именно равномерное их распределение. Отсутствие данных по равномерным распределениям можно было бы отнести за счет того, что многие исследователи выбирали более легкие для изучения, т. е. обычно сравнительно редко встречающиеся в сообществах, виды.
Однако в лесных сообществах Тринидада Грейг-Смкт, изучив все виды деревьев, обнаружил лишь крайне слабые следы равномерного распределения, даже после того как особи разных видов были сгруппированы в классы по размерам. Джонс, работавший в лесах Нигерии, также не упоминает о наличии равномерного распределения. Хотя относительную редкость равномерного распределения нельзя еще считать окончательно доказанной, при существующем положении вещей вполне оправдано сосредоточение внимания на групповых распределениях.
Если распределение является групповым, то независимо от причин, лежащих в основе этого явления, особи размещаются на поверхности пятнами; при этом либо встречаются пятна, где вид присутствует, и пятна, где он отсутствует, либо пятна, где он более обилен, и пятна с меньшим обилием. Если требуется выяснить причины этой пятнистости, желательны, а иногда существенно необходимы, сведения о размерах няъен. Это позволит по крайней мере исключить из числа возможных причин факторы, не обнаруживающие пятнистости того же масштаба.
Распределение особей может не только характеризоваться наличием пятен двух типов, но быть и еще более сложным; например, вид может образовывать группы особей, происходящие от одной с помощью вегетативного размножения, а эти группы могут быть более многочисленны в одних пятнах, чем в других. Другими словами, одновременно может существовать гетерогенность разного масштаба, и желательно бывает выявить существование этих разных типов неоднородности и их приблизительные размеры.
Некоторые указания масштабы неоднородности можно получить, меняя размеры учетной единицы при использовании описанных выше методов и отмечая, при каком размере отклонения от случайности перестают выявляться совсем или резко уменьшаются; однако это трудоемкий и малочувствительный способ. Во всяком случае, он имеет небольшое практическое значение для исследования неоднородности более значительных масштабов, обычно вызываемой изменчивостью факторов среды. Если неслучайность выражена достаточно резко, для выявления неоднородности относительно большого масштаба бывает достаточно произвести систематическое взятие образцов (например, с помощью сетки смежных квадратов) и после этого нанести на план изономы (т. е. линии, соединяющие точки с равной численностью или равными значениями других характеристик).
Грейг-Смит предложил более чувствительный метод, основанный на систематическом отборе образцов. Используется решетка смежных квадратов, число которых на каждой стороне решетки является какой-либо степенью 2, например, 16 X 16, 32 X 64. Число особей на каждый квадрат или другого типа ячейку решетки подсчитывается обычным путем. Общая дисперсия для всех ячеек решетки распределяется с помощью обычного метода дисперсионного анализа по группам из 2, 4, 8, 16 и т. д. ячеек, квадратным для четных степеней двух и удлиненным для нечетных, а остаток характеризует различия между отдельными ячейками в группах из двух ячеек, т. е. то, что можно назвать различиями «между группами из одной ячейки». Если расположение особей на всей изучаемой площади является вполне случайным, средний квадрат отклонений для групп всех размеров будет одинаков и равен среднему числу особей на одну ячейку сетки, т. е. отношение дисперсии к среднему будет равно 1 при любой величине образца.
Если распределение является групповым и растительный покров обнаруживает пятнистость, следует ожидать (Бартлетт, цит. по, что дисперсия будет возрастать при увеличении числа ячеек в группе вплоть до величины группы, соответствующей размеру пятен. Это подтверждается проверкой на модели сообщества, состоящего из кружков с заранее известным размещением. Если пятна сами распределяются случайно или образуют местные скопления, дисперсия при увеличении размеров группы будет сохраняться на прежнем уровне. Если пятна распределяются регулярно (что всегда наблюдается, когда на изучаемом участке чередуются пятна с большей и меньшей численностью и одинакового размера), дисперсия при дальнейшем увеличении размера группы будет уменьшаться. Если имеется несколько ступеней неоднородности, изменения дисперсии будут повторяться, когда группы ячеек решетки будут достигать соответствующих размеров. После проведения дисперсионного анализа полученных данных строится график зависимости дисперсии от размера групп ячеек и отмечаются наблюдаемые резкие подъемы и пики.
Проба на значимость представляет значительные трудности.
Когда этот метод был впервые предложен, сравнивали дисперсию при наибольших размерах группы квадратов и дисперсии групп, состоящих из одного квадрата, с помощью отношения дисперсий, или критерия F. В качестве модификации этого метода можно тем же способом сравнивать соседние точки кривой. Однако, как указал Бартлетт (личное сообщение), если доказано наличие неслучайного распределения, использование критерия F становится неправомерным. Томпсон, исследовавший эту проблему, показал, что, если строится некоторая правдоподобная (в математическом смысле) модель сообщества, ожидаемая дисперсия и ее стандартное отклонение могут быть рассчитаны для группы любого размера. Если наблюдаемые дисперсии не обнаруживают значимого отклонения, модель можно рассматривать как удовлетворительное описание типа размещения. Необходимость построения модели делает этот, несомненно гораздо более строгий, метод в большинстве случаев непрактичным.
Мы, таким образом, вынуждены оставаться на уровне субъективного суждения относительно пиков, хотя критерий F имеет «отрицательную ценность», выявляя различия, наверняка не являющиеся значимыми. На практике выявлению пиков существенно способствует их постоянство в серии анализов; если пик сохраняет то же самое положение или обнаруживает правильное смещение в ряду близких сообществ, сомнений в его реальности почти не остается. Кэршоу, проанализировав большое число искусственных сообществ, продемонстрировал общую применимость этого субъективного суждения. Остаточную дисперсию для групп из одного квадрата можно в случае необходимости проверить на отношение к среднему тем же способом, как и данные, полученные при случайном выборе площадок.
Отличие этого отношения от единицы имеет стандартное отклонение, ![]() где N — число степеней свободы, соответствующее группам из одного квадрата.
где N — число степеней свободы, соответствующее группам из одного квадрата.
Томпсон подтвердил возможность использования сходства графиков как основы для суждения о значимости.
Он показал также, что для случайного распределения можно легко рассчитать доверительные интервалы для любого требуемого уровня вероятности. Если распределение имеет дисперсию, равную единице, то сумма квадратов при N степенях свободы распределяется как Так, например, для 95%-ных доверительных интервалов верхний предел U находят по величине соответствующей Р — 0,025, а нижний предел L — по величине, соответствующей Р = 0,0975. Пределы для дисперсии в этом случае равны U/N и L/N. Расчет доверительных интервалов особенно полезен при небольшом числе анализов. Томпсон приводит в своей работе краткую таблицу 95%-ных пределов; более полная таблица из работы Грейг-Смита, содержащая практически все требуемые значения. Значения пределов выражены в единицах дисперсии, т. е. для данных по численности как отношение дисперсии к среднему значению.
Метод можно существенно усовершенствовать на основе предложения Кэршоу о применении одинакового подхода к изучению встречаемости и покрытия.
Вместо сетки квадратов используется некоторое количество параллельных рядов квадратов, и данные подсчета в каждом ряду объединяются последовательно в группы все большего размера. Пики на кривой зависимости дисперсии от размера группы дают в этом случае указание не на площадь пятен, а на их линейные характеристики. Это дает существенную экономию в числе учетных квадратов, требуемом для выявления гетерогенности любого масштаба. Далее, если есть основания предполагать, что пятна не являются изодиаметрическими, например на склонах, можно производить отдельные анализы в двух перпендикулярных направлениях. Кэршоу указывает, что дисперсию для наибольшего размера групп (полная длина трансекты) не следует принимать во внимание, так как она может частично зависеть от расстояния между рядами.
Вместо численности в качестве показателя неоднородности можно использовать встречаемость, если каждую клетку решетки разделить на более мелкие части, в каждой из которых отмечать присутствие или отсутствие вида; например, клетка может состоять из 25 мелких квадратов, так что можно отмечать любую суммарную величину от 0 до 25. Данные затем обрабатываются в точности так же, как данные по численности. Использование встречаемости имеет то преимущество, что можно проводить учет для любого вида, однако серьезным недостатком этого варианта метода является увеличение размеров ячеек решетки, что необходимо, если предполагается отдельно вести учет в более мелких делениях ячейки.
Кэршоу распространил применение метода на измерение покрытия, используя линейные трансекты, на которых в равномерно размещенных точках отмечается присутствие и отсутствие растений данного вида. Этот метод при размещении точек на расстоянии 1 см друг от друга оказался удовлетворительным для травянистых сообществ, к которым большинство других методов анализа размещения неприменимо. Основной единицей учета служат 5 последовательных точек на линии. Дисперсия для групп, состоящих из одной учетной единицы, при анализе покрытия, как и при анализе встречаемости, может быть подвергнута испытанию на сходство с дисперсией, ожидаемой в соответствии с биномиальным распределением с помощью критерия F (щ — число степеней свободы, соответствующее данному значению дисперсии, Сначала предполагалось, что такие данные должны быть перед анализом подвергнуты угловому преобразованию, однако дополнительное рассмотрение вопроса показало, что, если не нужно производить тест на значимость, такое преобразование не приносит существенной пользы. Если, однако, нужно получить доверительные интервалы, преобразование данных необходимо. Ожидаемая дисперсия для преобразованных данных приблизительно равна ![]() (где п — число точек или квадратов в основной учетной единице), но когда п мало, эта величина значительно отклоняется от истинной. Последнюю можно найти лишь путем расчета по соответствующему биномиальному разложению, куда вместо числа занятых точек подставляются преобразованные значения. Дисперсии для преобразованных данных при наиболее обычных значениях.
(где п — число точек или квадратов в основной учетной единице), но когда п мало, эта величина значительно отклоняется от истинной. Последнюю можно найти лишь путем расчета по соответствующему биномиальному разложению, куда вместо числа занятых точек подставляются преобразованные значения. Дисперсии для преобразованных данных при наиболее обычных значениях.
[1] Оно определяется двумя параметрами: I и m. Вероятность попадания участка территории х в пробную площадку рассчитывается по формуле: