Обмен веществ у автотрофных организмов
Организм — это открытая система, общность химических и энергетических превращений которой (обмен веществ — метаболизм) находится в равновесии с окружающей средой — динамическое равновесие.
Вещество окружающей среды может ассимилироваться в организме (обмен строительным материалом), из низко энергетических соединений синтезируются вещества с более высоким энергетическим потенциалом (ассимиляция, анаболизм). В процессе разложения высоко энергетических веществ (диссимиляция, катаболизм) освобождается энергия, которая используется для поддержания жизненных функций организма (например, функциональный обмен веществ: дыхание, брожение). Постоянный обмен веществ с окружающей средой происходит через поверхность тела и соответствующие органы.
В организме энзимы, часто в сочетании со вспомогательными веществами — кознзимами, воздействуют на вещества, определяя протекание необходимых реакций.
Благодаря объемной структуре и распределению зарядов в активном центре энзимы "распознают" молекулы, подлежащие разложению (специфические для субстрата), снижают необходимую начальную энергию (энергию активации) и "принимают решение" о необходимых изменениях субстрата (избирательное воздействие). Благодаря действию энзимов равновесие между исходными и конечными продуктами не смещается.
Если реакция идете выделением энергии (экзогенная реакция), то в присутствии специфических энзимов она протекает спонтанно
Если для получения конечного продукта необходимо затратить энергию (эндогенная реакция), то реакция не может ни начаться самопроизвольно, ни протекать даже в присутствии энзимов. Она может проходить только вместе с сильной экзогенной реакцией. Чаще всего это происходит посредством переноса остатков фосфорной кислоты или фосфатных групп АТФ (ГТФ, УТФ, КТФ) к менее энергоемкому реагенту, в результате чего он приобретает высокоэнергетическую связь. При расщеплении фосфатных групп высвобождается достаточно энергии, чтобы покрыть энергетический дефицит эндогенной реакции.

Рисунок. Цикл дикарбоновой кислоты растения С4.
Автотрофные организмы используют энергию солнечного света для синтеза энергоемких соединений (рисунок Цикл дикарбоновой кислоты растения С4):
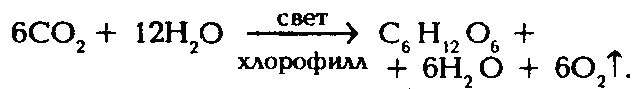
Фотосинтез протекает двумя взаимозависимыми парциальными реакциями:
- первичной, зависящей от света (световая реакция),
- вторичной, независимой от света (темновая реакция, рисунок В).

Рисунок. Зависимость фотосинтеза и дыхания от температуры.
В тилакоидных мембранах хлоропластов (рисунок — Зависимость фотосинтеза и дыхания от температуры, Разрез листа растения С4) с помощью пигментов, абсорбирующих свет, — до 400 молекул хлорофилла на светоприемник (рисунок Д) — электроны (е~) переносятся к молекулам-акцепторам, которые в фотосистеме 1 (ФС I) действуют как сильный восстановитель, а в фотосистеме II (ФС II) — как сильный окислитель.

Рисунок. Разрез листа растения С4.
Абсорбционный максимум фотосистемы I лежит в области 700 нм (Р 700) и в темноте обладает окислительно-восстановительным потенциалом + 0,4 В. Под действием светового возбуждения окислительно-восстановительный потенциал изменяется до -0,6 В. Этого достаточно, чтобы присоединить электроны к субстанции, восстанавливающей ферредоксин (ФВС — ферредоксино восстанавливающая субстанция), и запустить транспортную цепочку к НАДФ+ (рисунок Расход СО2 в зависимости от интенсивности солнечного излучения).

Рисунок. Расход СО2 в зависимости от интенсивности солнечного излучения.
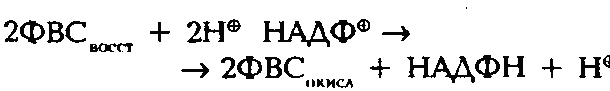
В фото стеме II с абсорбционным максимумом 680 нм (Р680) свет возбуждает сильное окислительное средство, которое в состоянии отобрать е- у Н2O с образованием O2 (фотолиз воды):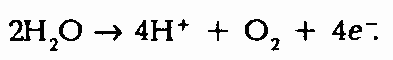
АТФ может быть синтезирован, если из фотосистемы 11 по транспортной цепочке электронов попадают в фотосистему 1 (нециклическое фотофосфорилирование), замещая недостающие в ней е-. Кроме этого, АТФ может появиться при циклическом потоке электронов в фотосистеме 1, если количество НАДФ+ недостаточно (циклическое фотофосфорилирование). Согласно Митчеллу (1961), благодаря протонному напору (электрохимическую разность потенциалов) между тилакоидным объемом и стромой при прохождении ионов через мембрану образуется АТФ.
Свет должен активировать неактивную форму синтеза АТФ, чтобы открылся какал Н+ и началось накопление АТФ. Темновая реакция (цикл Калвкга) требует наличия продуктов световой реакции НАДФН/Н+ и АТФ (рисунок Суточный кислотный цикл растений ММК) и может протекать без света, пока эти продукты есть в наличии.

Рисунок. Суточный кислотный цикл растений ММК.
О2 появляется здесь как промежуточный побочный продукт темновой реакции, протекающей в строме хлоропластов, если СO2 реагирует с рибулозодифосфатом с образованием 2 фосфорглицератов (молекулы с 3 атомами С, растения С3); при этом газообразный СО2 восстанавливается до исходных углеводов и фиксируется (рисунок Е). Для дальнейшей перестройки до фруктозы-6-фосфата, или глюкозы-б-фосфата АТФ поставляет энергию, а ионы НАДФН/Н+ — водород. Образовавшиеся АДФ + Р и НАДФ+ должны регенерироваться по световой реакции до АТФ и НАДФН/Н+.
Суммарная формула фотосинтеза:

Аппарат фотосинтеза может быть разрушен не только ядами из окружающей среды, но и под действием большинства гербицидов.
