Зависимость интенсивности обмена от совместного действия температуры и содержания кислорода в воде.
С повышением температуры при постоянной концентрации кислорода увеличиваются потребление кислорода и интенсивность обмена (рисунок 2), т. е. чем выше температура, тем при большем содержании кислорода меньше его потребление. Из данных Д. Фергюсона следует, что постепенное понижение содержания кислорода с 4 до 2 мг/л приводит к снижению интенсивности активного обмена в интервале температур от 5 до 25°С (рисунок 3). Значительный интерес представляют результаты опытов Д. Грехэма, проведенных на сеголетках Salvelinus fontinalis в лаборатории Ф. Фрая.
Изучая совместное действие температуры и уровня кислорода на интенсивность обмена, она выявила два интересных факта. Если уровень стандартного обмена постоянно возрастает по мере повышения температуры, то активный обмен увеличивается лишь при подъеме температуры до 20°С, а затем начинает падать, несмотря на дальнейшее повышение температуры.
Кроме того, интенсивность метаболизма рыб определяется температурой лишь при оптимальном содержании кислорода в воде.

Рисунок. 1. Зависимость летальной для карпа концентрации кислорода от температуры.
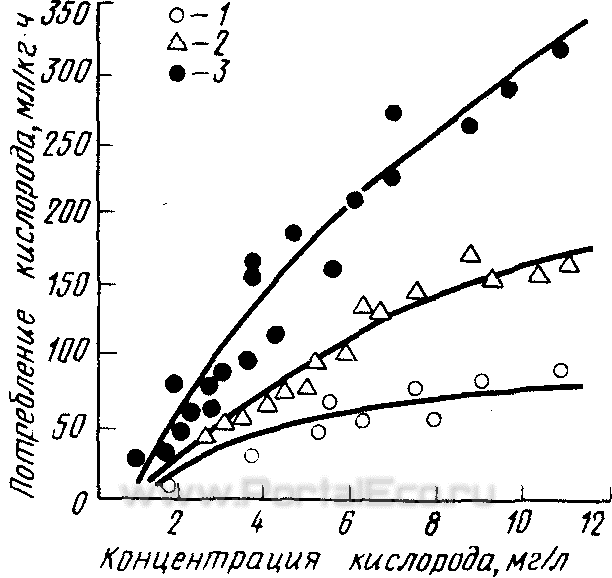
Рисунок. 2. Зависимость активного обмена Amphidon alosoides от температуры и концентрации кислорода.
1 — при температуре 5° С; 2 — при 10° С; 3 — при 15°С.
Если насыщение воды кислородом достигает критической величины и падает, то значение температурного фактора в определении уровня обмена полностью нивелируется и на первый план выступает содержание кислорода.
Эти факты имеют существенное значение для интерпретации результатов опытов, направленных на выявление зависимости уровня устойчивости рыб к ядам от содержания кислорода в воде.

Рисунок. 3. Совместное действие концентрации растворенного кислорода и температуры на потребление кислорода Perca flavescens.
Цифрами обозначены концентрации растворенного кислорода:
1 - воздушное насыщение; 2 - 4 мг/л; 3 - 3 мг/л; 4 - 2 мг/л; 5 - контроль.
При экспериментальном изучении общих и частных закономерностей интенсивности обмена у рыб, влиянии на него различных факторов водной среды, величины пороговой и критической концентраций кислорода важное значение имеет исходное физиологическое состояние подопытных рыб, определяемое многими факторами. Среди этих факторов первостепенное значение приобретает уровень кислорода, характерный для естественного водоема, из которого получены подопытные рыбы, длительность их акклимации к аквариальным условиям при том или ином кислородном режиме.
Это особенно важно при опытном установлении величин критической и пороговой концентраций кислорода для различных видов рыб, поскольку предварительная акклимация к уровню кислорода (пониженному или повышенному) влияет на кислородную устойчивость и чувствительность рыб. Первое обстоятельное изучение акклимации рыб к пониженному или повышенному содержанию кислорода в воде выполнено А. Г. Минц. Сеголетки подопытных рыб (карп, линь, карась, елец, окунь) длительное время находились в воде с пониженным (8 — 10%) или повышенным (120-130% 02) содержанием кислорода. Уже спустя несколько дней после начала опытов у рыб, находившихся при повышенном содержании кислорода, уровень обмена начинал возрастать, а у рыб, находившихся при пониженном содержании кислорода - снижаться.
Необходимо подчеркнуть, что определение интенсивности потребления кислорода подопытными рыбами проводилось при нормальном насыщении воды кислородом.
Новый постоянный уровень обмена устанавливается спустя 2 — 3 недели после начала акклимации, однако выраженность наступивших изменений была неодинакова у разных видов и определялась особенностями их экологии. Повышение интенсивности обмена при акклимации к повышенному содержанию кислорода у линей составило 55% по отношению к начальному уровню обмена, у окуней - 39%, у карпов 36%, у карасей 23,5%, у ельцов - 11%. При акклимации рыб к пониженному содержанию кислорода исследованные виды рыб как бы расположились в обратном порядке, т. е. снижение интенсивности обмена у ельцов составило 33%, у окуней — 17%, а у линей, карасей и карпов соответственно 15, 12 и 8% от исходного уровня обмена. Особого внимания заслуживает тот факт, что кислородная устойчивость акклимированных рыб, оцениваемая по величине пороговой концентрации, вызывающая удушье, изменяется в строгом соответствии с изменением уровня обмена.
Результаты исследований А. Г. Минц были подтверждены и другими специалистами, изучавшими этот вопрос на других видах рыб. Отметим прежде всего работу Н. Д. Никифорова, выполненную на молоди лосося, которая выращивалась в двух различающихся по кислородному режиму прудах. В одном из прудов содержание кислорода бы почти в два раза выше (10 — 12,5 мг/л), чем в другом пруду (5,5 мг/л). Интенсивность потребления кислорода у молоди из первого пруда оказалась значительно выше, чем у молоди из второго пруда.
Повышение способности некоторых видов лососевых рыб переносить низкие концентрации растворенного кислорода (1,8 — 2,4 мг/л) после выдерживания их в течение 4 недели при пониженном содержании кислорода в воде продемонстрировано также А. Д. Дэвисон. Активность рыб при этом несколько снижена, но угнетение может быть снято повышением содержания кислорода до 3 мг/л. Точно так же устойчивость ручьевой форели к дефициту кислорода может быть повышена путем предварительной адаптации к низким концентрациям кислорода.
Кислородный порог при этом снижается почти в два раза.
Устойчивость рыб повышается, а летальный порог снижается после предварительной адаптации к пониженным концентрациям кислорода. М. Шепард предполагает, что этот эффект обусловлен повышением способности рыб поглощать и связывать кислород при низких концентрациях, т. е. речь идет об улучшении транспортной функции крови и повышении сродства гемоглобина крови к кислороду. Это предположение подтверждается существованием связи между устойчивостью рыб к дефициту кислорода и сродством гемоглобина крови к кислороду: чем выше это сродство, тем выше устойчивость к недостатку кислорода и ниже кислородный порог. Однако можно дать и другое объяснение этому факту.
Повышение устойчивости рыб к дефициту кислорода после предварительной адаптации к пониженному кислородному режиму может быть обусловлено снижением уровня активного и стандартного обмена. В пользу этого предположения свидетельствуют результаты опытов А. Г. Минц.
