Повышение информативности селекционного процесса.
За последние годы интенсивно развивается биологическая кибернетика, изучающая специфические для живых существ общие принципы и конкретные механизмы целесообразного саморегулирования и активного взаимодействия с окружающей средой.
При этом главными являются три аспекта:
- самоорганизация биологических систем;
- информационные процессы;
- процессы управления (Коган и др., 1977).
Биологическая кибернетика изучает биологические системы разного уровня: от субклеточного до биосферного. Этапы развития биологической кибернетики отражены в ряде работ (Рашевский, 1968; Кастлер, 1968; Левине, 1968; Мак-Артур, 1968; К. Х. Уоддингтон и Р.Левонтин, 1970; Уотермен, 1971; Коган и др., 1977; Вильнер, Пешее, 1977; Антомонов, 1977; Моисеев, 1988 и др.). Нас будет интересовать возможность использования кибернетических подходов в эволюции и селекции.
Наиболее полно кибернетическое осмысление эволюционных процессов отражено в классических работах И. И. Шмальгаузена (Шмальгаузен, 1968, 1972). Им предложена общая схема регулирующего механизма эволюции (рисунок ).
При этом эволюция может рассматриваться как автоматически регулируемый процесс. И. И. Шмальгаузен выделяет два канала связи для передачи информации. Первый канал — реализация генетической информации в процессе онтогенеза (носитель прямой информации — ген). Второй канал — естественный отбор приспособленных генотипов в результате действия биоценоза на популяцию особей (носитель обратной информации — фенотип).

Общая схема регулирующего механизма эволюции (Шмальгаузен. 1968)
По мнению И. И. Шмальгаузена, в свете положений кибернетики гораздо ярче выдвигается различная роль обеих основных форм ее естественного отбора — стабилизирующей и движущей. Под действием движущей формы происходят усложнение фенотипа как носителя обратной информации и усложнение наследственного аппарата как механизма передачи прямой информации и основы его преобразования в индивидуальном развитии. Однако в прогрессивной эволюции организма как целого, в развитии его устойчивости ведущую роль играет стабилизирующая форма отбора. Через элиминацию всех случайных нарушений фенотипа она ведет к организации наиболее надежных каналов связи, к наибольшей помехоустойчивости как в передаче наследственной информации, так и в ее преобразовании в индивидуальном развитии и в реализации фенотипов как носителей обратной информации.
При передаче информации по первому и второму каналам связи могут возникать помехи, связанные с необычными, случайными факторами среды. Помехи в первом канале связи (радиация, температура, химические реагенты и др.) и во втором (хищники, паразиты, климатические аномалии) — всегда отрицательный фактор, ограничивающий не только жизнь и размножение популяции, но и возможность ее прогрессивной эволюции (Шмальгаузен, 1968). В одной из последних И. И. Шмальгаузен (1972) отмечает.
Что к конструктивным внешним факторам эволюции следует отнести большое количество информации в обоих каналах связи, означающее высокую мобильность популяции, а также высокое качество информации (системные связи), обеспечивающее устойчивость популяции в биогеоценозе.
При этом автор отмечает важную роль стабилизирующей формы естественного отбора для повышения устойчивости генотипов, популяций и всего эволюционного процесса в целом. Адаптивные реакции и стабилизация адаптивных форм обеспечивают устойчивость самой эволюции через организацию безошибочной передачи информации и ее трансформации.
Не менее важно использование кибернетического подхода в селекции, поскольку селекция, по Н. И. Вавилову (1987), представляет собой эволюцию, направляемую волей человека. Можно согласиться с М. А. Кадыровым (1991), назвавшим селекцию микроэволюционным процессом, сжатым во времени и пространстве. Последним в числе ряда методологических принципов оптимизации селекции автогамных растений сформулирован принцип достаточной информативности селекционного процесса (информация как одна из целей его функционирования). Автором разработаны система и технология информационного обеспечения управления селекционным процессом на базе персональных ЭВМ. Как справедливо отмечает М. А. Кадыров, приведение селекционного процесса для компьютеризации требует значительной системной проработки технологии селекции.
Следует отметить, что информационное обеспечение селекционного процесса не означает только использование современных методов генетического анализа или хранения громоздкой информации о селекционном материале в памяти ЭВМ. В первую очередь необходимо определить, какая информация нужна селекционеру. На наш взгляд, информационное обеспечение селекционного процесса должно решить следующие главные задачи:
Получение объективной эколого-генетической информации на каждом этапе селекции (в каждом питомнике). Адекватность этой информации идиотипу, т.е. создание единой информационной системы по принципу близости информации о селекционном материале и идиотипе (принцип "попадания в лузу"). Возможность контроля не только среднего значения признака, но и нормы реакции, поскольку наследуется не признак, а норма реакции. Возможность контроля не только генотипов по их фенотипам, но и среды как фона для отбора. Равноценность информации о генотипе и информации о среде. Решение проблемы дефицита эколого-генетической информации в различных узких местах селекционного процесса, в первую очередь в ранних поколениях F2 — F4. Возможность отбора по признаку на всем протяжении жизненного цикла растения.
Прежде всего решение этих задач невозможно без изменения представления об идиотипе как модели сорта. Традиционного представления об идиотипе как наборе количественных признаков явно недостаточно. Неполным будет идиотип, включающий требования к генотипу и условиям выращивания (Бороевич, 1981). Очень важной для построения идиотипа наряду с требованиями к генотипу, на наш взгляд, является экологическая ниша сорта как совокупность абиотических и биотических факторов среды, где сорт может обеспечивать высокую и устойчивую урожайность при хорошем качестве. Только в том случае будет реализован принцип экологической направленности селекционного процесса (Кильчевский, 1987).
Н. И. Вавилов (1987) выявил ряд географических закономерностей и районировании сортов пшеницы по территории бывшего СССР. Одни сорта занимали обширные территории, другие — определенные районы. Им было отмечено, что мы еще мало знаем природу космополитизма и узкой специализации не только сортов, но даже целых культур. Внимание селекционера преимущественно направляется на выведение сортов, на генотипические различия, на наследственную изменчивость. Чем глубже идет исследование, чем выше требования к селекции, тем больше приходится уделять внимания и лимоотношению среды и сорта, выявлению индивидуальных сортовых особенностей в смысле требования агротехники, удобрения, подбора определенных условий и районов культуры (Вавилов, 1987).
На наш взгляд, весьма продуктивным может быть использование кибернетических подходов к эволюции, разработанных И.И.Шмальга узеном (1968) применительно к информационному обеспечению селекционного процесса.
Поскольку действие естественного отбора имеет место и при селекции, сохраняют свое значение первый канал связи (н онтогенезе) и второй канал связи (на уровне популяции). Однако передача информации по первому и второму каналам связи имеет с мою специфику.
Специфика реализации генетической информации по первому каналу связи выражается в первую очередь в комфортных условиях культивирования (обработка почвы, удобрения, поливы, борьба с сорняками и др.) и управляемости онтогенезом по воле человека. В результате в культуре обычно сильно проявляются признаки, связанные с условиями культивирования и представляющие интерес для человека. Среда подставляет под действие отбора одни признаки и может не выявлять другие. При этом приспособительное значение признаков и их выражения при культивированиии отходят на второй план.
К. Х. Уоддингтон (1970) показал, что фенотип представляет собой эпигенетическое явление, функцию, зависящую от условий реализации генотипа и времени. Согласно идеям К. Х. Уоддингтона (1964, 1970), развивающийся организм выбирает один из нескольких возможных путей развития и эта единственная траектория онтогенеза (креод) характеризуется наибольшей устойчивостью. Совокупность всех возможных траекторий канализирована и образует эпигенетический ландшафт (рисунок 2). Поддержание на постоянном уровне процесса изменения (траектории) во времени автор назвал гомеорезом (стабилизированный поток) в отличие от гомеостаза (стабилизированное состояние). Наличие ряда траекторий является проявлением взаимодействия генотипхсреда на уровне онтогенеза особи.
При этом селекционера в первую очередь будут интересовать два момента:
- оптимальные условия культивирования, в которых проявляется признак;
- оптимальная стадия онтогенеза, на которой возможен отбор, в том числе косвенный.
Что касается первого момента, здесь наиболее важным является типичность условий культивирования особей в данной среде по отношению к будущим условиям возделывания.
Второй момент имеет принципиальное значение для селекции ряда признаков, в первую очередь определяющих устойчивость. Многочисленны работы физиологов для создания экспресс-методов отбора на устойчивость к ряду факторов по проросткам и вегетирующим растениям. За последние годы интенсивно развивается клеточная селекция, позволяющая вести отбор уникальных генотипов в культуре in vitro. Новым направлением в селекции является гаметная и зиготная селекция — отбор на уровне гаметофита и эмбриональных клеток. Все эти методики требуют отпета на один принципиальный вопрос — есть ли связь между проявлением признака на любой стадии онтогенеза и его проявлением на уровне взрослого растения. Для решения этой проблемы необходим ряд методических экспериментов, чтобы "прозвонить" всю цепочку от гамет до взрослого организма, весь первый канал связи и установить оптимальную и возможные фазы для отбора по изучаемому признаку или некоторым косвенным признакам. Отдельные аспекты данной проблемы обсуждаются нами в последующих главах.
Второй канал связи, выявленный И. И. Шмальгаузеном, присутствует и в селекционном процессе.
Он также имеет свою специфику. Специфика эта проявляется в ослаблении действия биогеоценоза на селектируемую популяцию (применение удобрений, пестицидов, орошения и др.). Главная же особенность отбора фенотипов в селекции в отличие от естественного заключается в организованности всех этапов селекционной работы, качественном отличии методов отбора (индивидуальный отбор, испытание по повторностям, испытание генотипов в нескольких средах и т.д.) в питомниках, в силу чего этот информационный канал в каждом поколении и в каждом питомнике будет иметь свою специфику. На наш взгляд, в связи с этим в селекции правильнее рассматривать не отдельные звенья селекционной цепи в качестве элементарных информационных каналов, а всю их совокупность, начиная от выбора исходного материала до использования сорта в конкретном регионе возделывания. Такой канал связи мы назвали большим информационным каналом, а реализацию генетической информации в онтогенезе — малым информационным каналом (Кильчевский, 1995). Введение этих терминов позволяет более четко подойти к проблеме использования информации в селекции.
Понятие "большой информационный канал" позволяет более конкретно поставить вопрос об адекватности эколого-генетической информации идиотипу, условиям культивирования в производстве, типизации условий отбора на всех этапах селекции и их привязке к будущим условиям возделывания, правильной экологической организации селекции и обеспечению ее экологической целенаправленности на конкретную эконишу сорта, отбору как по среднему значению признака, так и по норме реакции. Потеря или искажение информации в большом информационном канале равноценна потере селекционного материала и снижению эффективности селекции.

Эпигенетический ландшафт — возможные траектории развития (Waddinglon. 1966)
Таблица Фенотипы девяти генотипов с различной нормой реакции на два фактора среды
|
Генотип |
Сочетание < |
>факторов среды |
|||||||||||||||
|
1 |
2 |
1 |
2 |
1 |
2 |
1 |
2 |
1 |
2 |
1 |
2 |
1 |
2 |
1 |
2 |
1 |
2 |
|
+ |
+ |
+ |
0 |
+ |
- |
0 |
+ |
0 |
0 |
0 |
- |
- |
+ |
- |
0 |
- |
- |
|
A D |
+ 1 4-1 |
+1 0 |
+1 -1 |
0 +1 |
0 0 |
0 -1 . |
-1 +1 |
-1 0 |
-1 -1 |
||||||||
|
А Е |
+ 1 0 |
+1 0 |
+1 0 |
0 0 |
0 0 |
0 0 |
-1 0 |
-1 0 |
-1 0 |
||||||||
|
A F |
+ 1 -1 |
+1 0 |
+1 +1 |
0 -1 |
0 0 |
0 +1 |
-1 -1 |
-1 0 |
-1 + |
||||||||
|
В D |
0 +1 |
0 0 |
0 -1 |
0 +1 |
0 0 |
0 -1 |
0 +1 |
0 0 |
0 -1 |
||||||||
|
В Е |
0 0 |
0 0 |
0 0 |
0 0 |
0 0 |
0 0 |
0 0 |
0 0 |
0 0 |
||||||||
|
В F |
0 -1 |
0 0 |
0 +1 |
0 -1 |
0 0 |
0 +1 |
0 -1 |
0 0 |
0 +1 |
||||||||
|
С D |
-I +1 |
-1 0 |
-1 -1 |
0 +1 |
0 0 |
0 -1 |
+1 +1 |
+1 0 |
+1 -1 |
||||||||
|
С Е |
-1 0 |
-1 0 |
-1 0 |
0 0 |
0 0 |
0 0 |
+1 0 |
+ 1 0 |
+1 0 |
||||||||
|
С F |
-L -1 |
-1 0 |
-1 +1 |
0 -1 |
0 0 |
0 +1 |
+1 -1 |
+1 0 |
+1 +1 |
Примечание: + 0 — средовая реакция генотипа по каждому фактору; +1 — лучшие фенотипы.
Большой интерес представляет вопрос о помехах в большом и малом информационном каналах и их устранении.
Главной помехой получения объективной информации является взаимодействие генотип — среда (ВГС). Конкретными причинами проявления ВГС могут быть резкие отклонения условий культивирования от нормальных (лимиты абиотических факторов, эпифитотии и др.). Генетической основой ВГС является выявленное В. А. Драгавцевым (1984, 1993) явление переопределения генетической организации количественных признаков. Автором показано, что при смене лимитирующих факторов может происходить явление смены спектра локусов, детерминирующих развитие признака, а также явление смены спектра модулей — элементарных единиц описания организации системы количественного признака. Каждый модуль состоит из трех взаимосвязанных признаков: результирующего и двух компонентных. В. А. Драглвцев пришел к выводу, что переопределение генетической организации признака проявляется на всех уровнях организаций живой материи, в том числе на организменном, популяционном и ценоти Чечком уровнях. Организменный уровень рассмотрен нами ранее, но пому кратко остановимся на популяционном (второй канал связи). Под действием ВГС среда "подставляет" под действие отбора ту или иную группу генотипов в популяции (в том случае, если генотипы Отличаются по норме реакции). Проиллюстрируем это на простом примере. Пусть имеются три генотипа А, В и С, отличающиеся положительной ( + 1), нейтральной (0) или отрицательной (-1) реакций на действие фактора среды 1. Тогда различия между генотипами в ргдах можно записать в виде простой матрицы
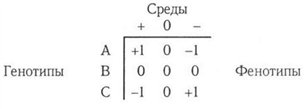
Аналогичным образом можно проинтерпретировать реакцию генотипов Л. Е и F на фактор 2. Более сложной будет модель, если Гепотипы отличаются по реакции на два фактора среды (таблица 1).
В этом случае возможны девять сочетаний генотипов по нормам реакции и девять сочетаний сред.
Заметим, что в каждой среде лучшим будет свой набор генотипов. Среда "подставляет" под отбор iy или иную группу особей, и только в средней среде (00) фактически не происходит отбор по норме реакции. Если увеличить число мчютипов, признаков, норм реакции, то число возможных вариантов лучших генотипов в средах неизмеримо возрастет. В принципе можно говорить об уникальности каждой среды как сочетания комплекса факторов при различном их значении (температура, свет, удобрения и др.). Все вышесказанное — достаточно простая иллюстрация ВГС ни популяционном уровне, однако позволяющая конкретизировать проблему типизации сред в большом информационном канале.
Если и регионе будущего возделывания сорта (эконише сорта) преобладает какое-либо сочетание факторов среды 1 и 2 (например, варианты +0, 0 0), очевидно, что на всех этапах селекции также должно быть создано преобладание этих факторов среды, в противном случае в результате ВГС какой-либо из этапов селекции направит изменчивость в популяции в нежелательную сторону. Целесообразно использовать биологические индикаторы — сорта-тестеры, которые, сохраняя или изменяя ранги в различных условиях среды в большом информационном канале под действием комплекса разнонаправленных факторов, могут служить свидетельством наличия или отсутствия помех. Резкая смена рангов сортов-индикаторов в сравнении с их поведением в будущей эконише сорта будет говорить о сильном проявлении ВГС и наличии помех в информационном канале. В этом случае следует снижать интенсивность отбора. Образно говоря, необходима настройка всех передатчиков информации на любом этапе селекции на волну принимающей станции (средовые и агротехнические условия возделывания сорта в предполагаемом ареале районирования).
Таким образом, нами предлагается использовать в селекции термины "малый информационный канал" — реализация генетической информации в онтогенезе и "большой информационный канал" — передача генетической информации при отборе в селектируемой популяции на всех этапах селекции. Повысить эффективность использования информации в обоих каналах и результативность селекции можно путем экологической организации селекционного процесса, адекватности (типичности) условий на каждом этапе селекции будущей эконише сорта, объективной информации как о генотипах (в том числе норме их реакции), так и о средах отбора, снятии помех, связанных со взаимодействием генотип — среда.
