Наследование каллусогенеза и регенерации у томата.
Дисперсионный анализ комбинационной способности генотипов по признакам "масса каллуса" и "регенерационщая способность" показал высокую достоверность различий по эффектам ОКС и СКС (таблица ). Соотношение средних квадратов ОКС/СКС по массе каллуса колебалось от 0,68 до 3,99; по регенерационной способности — от 1,11 до 2,68, что свидетельствует о преобладании аддитивных эффектов над неаддитивными. Тем не менее сопоставление отношений средних квадратов ОКС/СКС по признакам in vivo и in vitro говорит о том, что в генетической детерминации последних неаддитивные эффекты играют несколько большую роль.

Каллусогенез у томата ( Талалихин, а - Доходный. 0 - Талалихин х Доходный)

Регенерация у томата (1 - Талалихин. 2 - Доходный. 3 - Талалихин Доходный Z7,) на различных средах: а — ИУК. 6 БАП. 6 - ИУК, кинетин
Таблица 31. Дисперсионный анализ комбинационной способности образцов томата по признакам "масса каллуса" и "регенерационная способность" в различных условиях среды.
|
Гормоны |
Условия культивирования |
Средние квадраты |
||
|
ОКС |
СКС |
ОКС/СКС |
||
|
|
Масса каллуса, мг |
|
||
|
6 БАП, 2.4-Д |
24—26° |
45562,7** |
22036,5** |
2,07 |
|
6 БАП, 2.4-Д |
11—13° |
10977,7** |
4684.4** |
2,34 |
|
6 БАП, НУК |
24—26° |
20161,5** |
29780.7** |
0,68 |
|
6 БАП. НУК |
11—13° |
110934,2** |
27780.8** |
3.99 |
|
|
Регенерационная способность, % |
|
||
|
ИУК, кинетин |
5—6 тыс.люкс |
341,99** |
308,90** |
1.11 |
|
ИУК, кинетин |
1—2 тыс.люкс |
160,76** |
114,21** |
1.41 |
|
ИУК. 6 БАП |
5—6 тыс.люкс |
124,91** |
94,24** |
1,33 |
|
ИУК, 6 БАП |
1—2 тыс.люкс |
196,09** |
73.09** |
2.68 |
Оценка параметров комбинационной способности генотипов томата в четырех средах (таблица) позволяет выделить по эффектам ОКС Доходный, Beta и Линию 7. Хуже всех на большинстве сред процесс образования каллуса проходил у S.a.mini. Все остальные генотипы меняли ранги в зависимости от температуры и гормонального состава среды. Вариансы СКС, как правило, были выше на средах, содержащих НУК. Наибольшими вариансами СКС обладали генотипы Доходный и Beta. Они могут быть использованы при гибридизации для повышения эффективности процесса каллусо-образования у томата.
Параметры комбинационной способности генотипов по регенера-ционной способности представлены в таблице. Характерно, что ни один из генотипов не сохранил всех положительных знаков эффектов ОКС на всех средах. К числу лучших может быть отнесена Линия 7. которая на среде с кинетином оказалась в числе средних, а на среде с 6 БАП — абсолютно лучшей. Остальные генотипы сильно изменяют ранги в зависимости от гормонального состава среды и условий освещенности. Так, например, сорт Талалихин имеет наименьший эффект ОКС на среде с кинетином при нормальной освещенности и наибольший — на среде при пониженной освещенности. Этот же сорт выделяется по вариансе СКС. Таким образом, процессы каллу-сообразования и регенерации сортоспецифичны, в значительной степени зависят от гормонального состава питательной среды и условий культивирования.
Таблица 32. Параметры комбинационной способности генотипов томата по признаку "масса каллуса" (в мг) в различных условиях среды.
|
Генотипы |
БАП. 2,4-Д |
6 БАП. НУК |
||||||
|
24-26‘С |
11—13С |
24-26 С |
11—13‘С |
|||||
|
оке |
скс |
оке |
скс |
оке |
скс |
оке |
скс |
|
|
Талалихин |
-10,6 |
16427.6 |
-23,8 |
5043,9 |
2,0 |
30889,9 |
-130,4 |
19307.2 |
|
Доходный |
86,2 |
32141,8 |
6.4 |
2843,5 |
-8,1 |
31871,8 |
87,3 |
26887.8 |
|
П-7 |
-72.1 |
26484,8 |
22,4 |
2529,1 |
-39.8 |
6936,8 |
-3.9 |
12081.2 |
|
Beta |
47.3 |
26370,8 |
50,7 |
4719.9 |
47,2 |
15672,3 |
59.8 |
22323,0 |
|
S.a.mini |
-115,2 |
9999,9 |
-54,9 |
3076,9 |
-87.8 |
56514,6 |
-89.6 |
14031,6 |
|
Линия 7 |
-13,6 |
9547,4 |
21.3 |
6113,4 |
31,7 |
14385,9 |
186,5 |
38274,2 |
|
L.pimpinellifolium |
24,6 |
22823.3 |
-23.0 |
3851,9 |
29,1 |
6119,2 |
-72,3 |
23784,1 |
|
Torosa |
53,4 |
16560,6 |
0.78 |
4516.2 |
25,8 |
13773,9 |
-37.3 |
44277,4 |
|
НСР05 |
37.4 |
— |
14.6 |
— |
76,2 |
— |
32.3 |
— |
Примечание. ОКС — константа ОКС; СКС — варианса СКС.
Таблица 33. Параметры комбинационной способности генотипов томата по признаку "регенерационная способность" (в %) в различных условиях среды.
|
Генотипы |
ИУК, кинетин |
ИУК, 6 БАП |
||||||
|
5—6 тыс. люкс |
1—2 тыс. люкс |
5—6 тыс. люкс |
1 —2 тыс. люкс |
|||||
|
ОКС |
СКС |
ОКС |
СКС |
ОКС |
СКС |
ОКС |
СКС |
|
|
Талалихин |
-6,19 |
576,9 |
6,72 |
149,6 |
4,60 |
194,5 |
1,65 |
126,2 |
|
Доходный |
12,30 |
218,9 |
-2,34 |
244,6 |
1,03 |
49.9 |
-10,40 |
86.9 |
|
П-7 |
-4,29 |
400,2 |
5,52 |
109,2 |
-3,35 |
28.9 |
0,62 |
50,9 |
|
Beta |
3.37 |
222.7 |
-2,44 |
66,8 |
-3,70 |
89.2 |
0.87 |
18.9 |
|
S.a.mini |
0,67 |
91,6 |
-3,10 |
83,9 |
-1,26 |
102.9 |
-1,19 |
58,9 |
|
Линия 7 |
0,24 |
187,5 |
-0,71 |
115,6 |
5,73 |
40.8 |
3.46 |
34,3 |
|
L.pimpinellifolium |
-3,72 |
277.6 |
-3,90 |
64,2 |
-2,28 |
25,0 |
2,35 |
49.0 |
|
Torosa |
-2,39 |
186.9 |
0,25 |
13,7 |
-0,79 |
191.3 |
2,60 |
47,6 |
|
НСР05 |
2.51 |
— |
1,28 |
— |
1,37 |
— |
1.31 |
— |
Примечание. ОКС — константа ОКС: СКС — варианса СКС.
Таблица Степень доминирования по признакам "масса каллус а" и "регенерационная способность"
|
Условия культивирования |
Гормоны |
Гибриды |
Hp>1 |
-1Hp⩽1 |
Hp<-| |
|
Масса каллуса, мг |
|||||
|
24—26° |
6 БАП. 2.4-Д |
ШТ. |
8(0) |
9(0) |
11(4) |
|
|
|
% |
28.6(0) |
32.1(21.4) |
39.3(14.3) |
|
|
6 БАП. НУК |
шт. |
18 |
7 |
3 |
|
|
|
% |
64.3 |
25,0 |
10.7 |
|
11—13° |
6 БАП. 2.4-Д |
шт. |
3 |
12 |
13 |
|
|
|
% |
10.7 |
42,9 |
46,4 |
|
|
6 БАП. НУК |
шт. |
13(6) |
7(5) |
8(4) |
|
|
|
% |
46.4(21.4) |
25.0(17.8) |
28.6(14.3) |
|
Регенерационная способность, % |
|||||
|
5—6 тыс.люкс |
ИУК. кинетин |
шт. |
10 |
9(8) |
9(3) |
|
|
|
% |
35.7 |
32,1(28,6) |
32.1(10.7) |
|
|
ИУК. 6 БАП |
шт. |
12(7) |
12(8) |
4(0) |
|
|
|
% |
42.9(25,0) |
42,9(28,6) |
14.3(0) |
|
1—2 тыс.люкс |
ИУК. кинетин |
шт. |
15(5) |
Ю(5) |
3(0) |
|
|
|
% |
53.6(17.9) |
35,7(17,9) |
10.7(0) |
|
|
ИУК. 6 БАП |
шт. |
7 |
10 |
11 |
|
|
|
% |
25 |
35,7 |
39,3 |
Примечание. В скобках число гибридов, соответствующих ограничениям Хеймана.
При различных условиях культивирования изменяется и характер наследования признаков in vitro (таблица ). Так, под действием НУК число гибридов с положительным сверхдоминированием по массе каллуса было преобладающим (46,4 — 64,3%), а под действием 2,4-Д чаще проявлялось отрицательное сверхдоминирование (39,3 — 46,4%). На двух средах из четырех имел место эпистаз, так как ряд линий был удален из диаллельной таблицы в связи с несоответствием ограничениям Хеймана.
Близкая картина по изменению характера наследования признака в зависимости от гормонального состава и освещенности наблюдалась и по регенерационной способности.
На трех средах из четырех чаще проявлялось положительное сверхдоминирование (35,7 — 53,6%), на четвертой среде (пониженная освещенность, ИУК + 6 БАП) чаще имело место отрицательное сверхдоминирование (39,3%). На трех средах проявлялся эпистаз.
Таблица 35 Параметры Хеймана по признакам "масса каллуса" и "регенерационная способность".
|
Условия культивирования |
Гормоны |
H1 D |
н2 4H1 |
JtDHi + F J4DH{ -F |
r |
Dmax |
Rmax |
h1 H1 |
|
Масса каллуса, мг |
||||||||
|
24—26°С |
6 БАП, 2,4-Д |
0,94 |
0.19 |
1.34 |
0,27 |
734.1 |
997,4 |
0.94 |
|
|
6 БАП. НУК |
1.77 |
0,19 |
2,67 |
-0,85 |
1443,4 |
646,9 |
0.88 |
|
11—13< |
6 БАП. 2,4-Д |
1,65 |
0,19 |
1,95 |
-0,56 |
355,6 |
30,8 |
0,31 |
|
|
6 БАП, НУК |
1.81 |
0,19 |
0,94 |
-0,94 |
557,2 |
155,4 |
0,15 |
|
Регенерационная способность, % |
||||||||
|
5—6 тыс люкс |
ИУК, кинетин |
1,64 |
0.22 |
1.71 |
-0,50 |
78.7 |
9,9 |
0.29 |
|
|
ИУК. 6 БАП |
0,98 |
0,23 |
1,63 |
0,53 |
45,2 |
97,3 |
0.29 |
|
1—2 тыс люкс |
ИУК. кинетин |
1.34 |
0,20 |
1,02 |
-0,84 |
67.9 |
36.4 |
1.36 |
|
|
ИУК. 6 БАП |
1.81 |
0,15 |
2,45 |
-0,23 |
66,9 |
45,0 |
0 |
Таким образом, комбинационная способность генотипов томата в культуре in vitro в сильной степени подвержена действию условий культивирования и в особенности гормонального состава среды. Оптимум средовых условий зависит от особенностей генотипа, в значительной степени варьирует.
Генетические параметры Хеймана по признакам "масса каллуса" и "регенерационная способность" представлены в таблице 35. По массе каллуса средняя степень доминирования варьирует от 0,94 до 1,81, что говорит о наследовании признака по типу неполного доминирования или сверхдоминирования. На всех средах, за исключением среды с НУК при пониженной температуре, преобладает действие доминантных генов. Коэффициент корреляции свидетельствует о смене направления доминирования. На среде с 2,4-Д при оптимальной температуре доминирует меньшая масса каллуса, в остальных условиях культивирования — большая масса каллуса. Это же подтверждается значениями полностью доминантного и полностью рецессивного родителя. Различия между генотипами детерминируются одним геном.
Расположение генотипов на графиках Хеймана (рисунок) дает возможность выявить проявление доминантных и рецессивных аллелей, действующих на признак.
Анализ графиков позволяет установить, что под действием условий среды может происходить изменение проявления действия аллелей. При этом имеют место сложные взаимоотношения типа "ген х гормон х условия культивирования". Так, например, сорта Талалихин и П-7 на среде с 2,4-Д при оптимальной температуре и на среде с НУК при пониженной температуре попадают на участок графика с преобладающим действием рецессивных аллелей, а в остальных условиях среды в этих генотипах действуют доминантные аллели. Практически не происходит изменение проявления действия аллелей в различных условиях среды у сорта Torosa, который является носителем доминантных генов. У Линии 7 и L.pimpinellifolium под действием пониженной температуры усиливается проявление рецессивных аллелей. Аналогичная картина наблюдается у S.a.mini под действием смены ауксина (2,4-Д — НУК). На средах с 2,4-Д при оптимальной температуре и с НУК при пониженной температуре проявляется эпистаз, что приводит к потере отдельных линий на графике Хеймана.

Генетика признака "масса каллуса"
Параметры Хеймана по признаку "регенерационная способность" свидетельствуют о проявлении полного или неполного доминирования. Произведение частот доминантных и рецессивных генов изменяется от 0,15 до 0,23. Как правило, преобладают доминантные гены, поскольку соотношение их с рецессивными колеблется от 1,02 до 2,45. Доминирование на трех средах из четырех направлено в сторону усиления регенерационной способности, а на среде с 6 БАП при нормальной освещенности — в сторону ослабления. Различия между генотипами, по всей видимости, обусловлены одним геном.
Анализ графиков Хеймана (рисунок 12) также свидетельствует об изменении проявления действия аллелей в различных условиях среды. Так, например, на среде с кинетином при нормальной освещенности образец П-7 расположен в средней части графика (равное действие доминантных и рецессивных аллелей), а на остальных средах — в нижней (действие доминантных аллелей). Тем не менее большинство линий расположены в левой части графиков, что свидетельствует о преобладании доминантных генов. На среде с 6 БАП при нормальной освещенности проявляется полное доминирование, в остальных средах — сверхдоминирование.
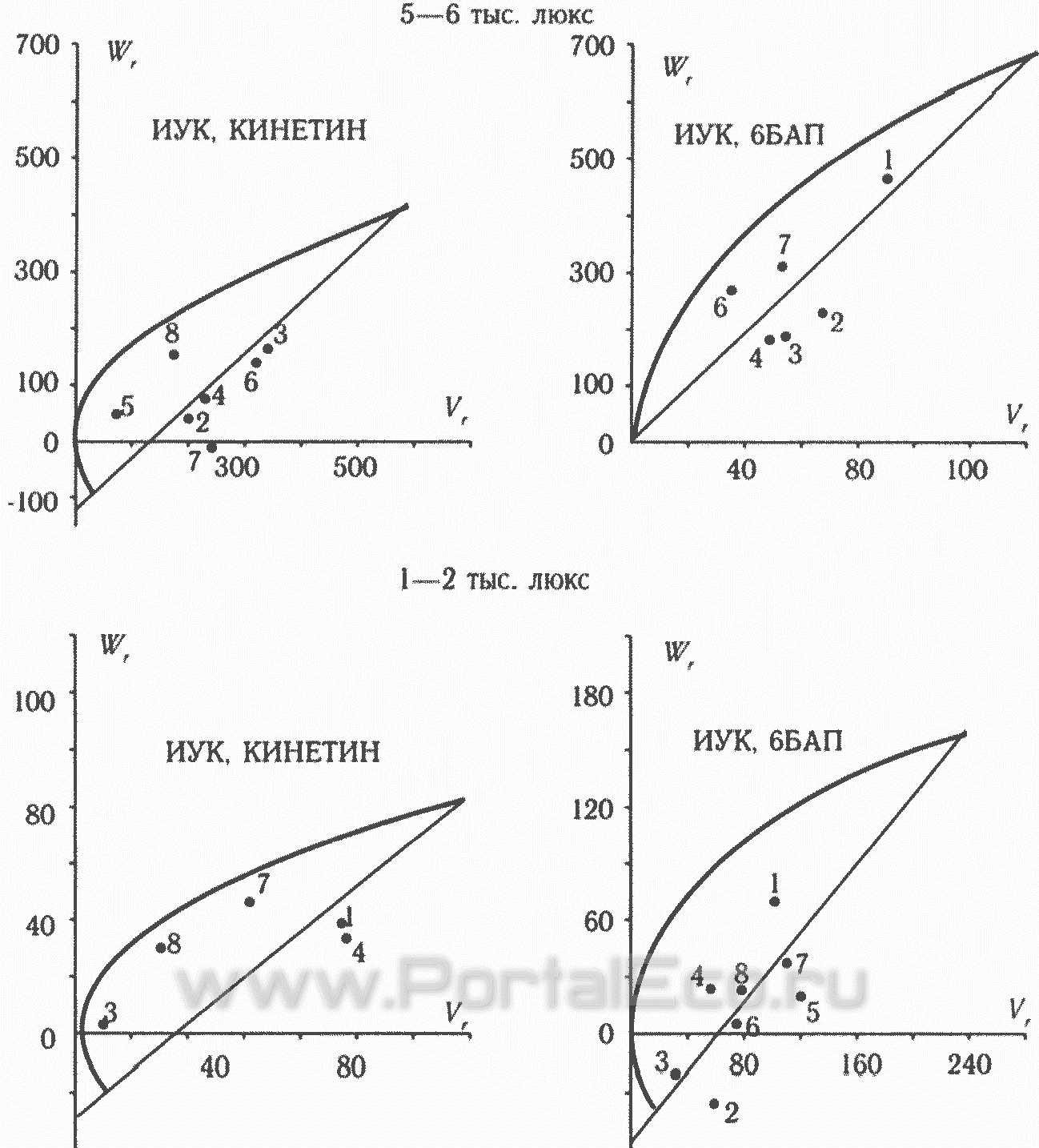
Генетика признака "регенерационная способность"
Таблица Среднее значение и относительная стабильность генотипов томата по признаку "масса каллуса" (в мг).
|
Номер генотипа |
Номер генотипа |
|||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
2 |
8 |
|
|
xi |
||||||||
|
1 |
759,3 |
633,2 |
610,1 |
756,0 |
643,1 |
555,2 |
636,6 |
615,2 |
|
2 |
|
733,5 |
665,7 |
745,1 |
849,4 |
861.6 |
851.1 |
731,2 |
|
3 |
|
|
624,4 |
755,0 |
688,9 |
767.8 |
659,0 |
746,1 |
|
4 |
|
|
|
732,3 |
699,8 |
904.7 |
753,3 |
808,8 |
|
5 |
|
|
|
|
422,0 |
667,1 |
534,5 |
579,2 |
|
6 |
|
|
|
|
|
741,9 |
805,3 |
894,3 |
|
7 |
|
|
|
|
|
|
697.2 |
636,8 |
|
8 |
|
|
|
|
|
|
|
735,0 |
|
Sgi |
||||||||
|
1 |
71,6 |
64,2 |
92,3 |
83,2 |
100,2 |
87.3 |
91.6 |
78,7 |
|
2 |
|
49,9 |
61,1 |
71,6 |
67,1 |
63.9 |
71.2 |
89,3 |
|
3 |
|
|
72,4 |
61,6 |
74,6 |
56,4 |
76,8 |
69,6 |
|
4 |
|
|
|
57,1 |
77.8 |
57.6 |
66.0 |
66,2 |
|
5 |
|
|
|
|
75,4 |
77.8 |
99,4 |
85,5 |
|
6 |
|
|
|
|
|
61.6 |
54,8 |
53,5 |
|
7 |
|
|
|
|
|
|
83.4 |
104.0 |
|
8 |
|
|
|
|
|
|
|
70.6 |
Судя по графикам, трудно идентифицировать генотипы, которые были бы бесспорными донорами высокой регенерационной способности. Проявление этого признака в сильной степени зависит от гормонального состава среды и условий культивирования.
В таблице представлены усредненные по четырем средам данные о массе каллуса у восьми родителей и гибридов между ними. Наибольшая масса каллуса отмечена у образцов Талалихин, Линия 7, Beta, Torosa и Доходный. Самое слабое каллусообразование — у формы S.a.mini. У гибридов чаще всего проявлялось положительное сверхдоминирование (13 гибридов), в 8 случаях отмечено промежуточное наследование и в 7 — отрицательное сверхдоминирование. По стабильности среди лучших форм выделились Доходный, Beta и Линия 7. В большинстве случаев (50%) имеет место положительное сверхдоминирование в сторону увеличения s (уменьшения стабильности) и только у 4 гибридов — отрицательное сверхдоминирование. Все эти четыре комбинации проявляют гетерозис по массе каллуса.
Таблица Среднее значение и относительная стабильность генотипов томата по признаку "регенерационная способность" (в %)
|
Номер генотипа |
Номер генотипа |
|||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
|
xi |
||||||||
|
1 |
63,4 |
69,7 |
68,8 |
64,3 |
65,5 |
61,2 |
53,9 |
45.9 |
|
2 |
|
60,8 |
57,3 |
48,5 |
54,4 |
66,3 |
59,7 |
63,2 |
|
3 |
|
|
51,8 |
57.4 |
63,3 |
63,5 |
63,9 |
57,6 |
|
4 |
|
|
|
57.8 |
55,2 |
67.2 |
62,0 |
64,1 |
|
5 |
|
|
|
|
52,0 |
64.6 |
59,7 |
60,3 |
|
6 |
|
|
|
|
|
62,9 |
52,7 |
59,6 |
|
7 |
|
|
|
|
|
|
54,0 |
60.4 |
|
8 |
|
|
|
|
|
|
|
63.7 |
|
Sgi |
||||||||
|
1 |
29,3 |
21,7 |
8,65 |
9,7 |
31,6 |
33,6 |
43,9 |
45,7 |
|
2 |
|
22,1 |
31,0 |
24.0 |
27,1 |
24,4 |
27,1 |
23,0 |
|
3 |
|
|
40,9 |
23,8 |
4.6 |
6,5 |
13,9 |
10.1 |
|
4 |
|
|
|
12,6 |
29,6 |
18,3 |
10,2 |
15.6 |
|
5 |
|
|
|
|
21,6 |
8.5 |
7.9 |
15.4 |
|
6 |
|
|
|
|
|
23,3 |
37.3 |
22.7 |
|
7 |
|
|
|
|
|
|
23,5 |
20,2 |
|
8 |
|
|
|
|
|
|
|
14,4 |
Различия между родителями по регенерационной способности (таблица 37) не столь велики, как по массе каллуса. Лучшими в среднем по всем средам являются Талалихин, Доходный, Линия 7, Torosa и Beta, худшими — П-7 и S.a.mini. В 13 случаях отмечено положительное сверхдоминирование, в 6 — отрицательное, в 9 — промежуточное наследование. Наиболее стабильными среди лучших форм были Beta и Torosa.
По стабильности в 11 случаях наблюдалось положительное сверхдоминирование, в 10 — отрицательное, в 7 — промежуточное наследование. У 9 гибридов гетерозис по регенерационной способности сочетался с гетерозисом по стабильности. Таким образом, намечается некоторый параллелизм в проявлении каллусогенеза и регенерации у родительских форм и их гибридов. К числу лучших по этим признакам можно отнести Талалихин, Доходный, Beta, Линия 7, Torosa. По обоим признакам в 13 случаях из 28 имеет место гетерозис, причем большинство гетерозисных гибридов отличаются большей стабильностью в сравнении с родителями. Признак "масса каллуса" подвержен
Таблица Параметры среды как фона для отбора геютипов томата по массе каллуса и регенерационной способности.
|
Условия культивирования |
Гормоны |
dk |
Ser |
tk |
Pk |
|
|
Масса каллуса, мг |
|
|
||
|
24—26”С |
6 БАП, 2,4-Д |
123,6 |
19,1 |
0.569 |
0.109 |
|
|
6 БАП. НУК |
643,2 |
12,0 |
0.707 |
0.085 |
|
11—13°С |
6 БАП, 2,4-Д |
-495,3 |
31,5 |
0.616 |
0,194 |
|
|
6 БАП, НУК |
-271.5 |
47,5 |
0.750 |
0.356 |
Регенерационная способность, %.
|
5—6 тыс.люкс |
ИУК, кинетин ИУК. 6 БАП |
-3.0 6.4 |
31,1 15,0 |
0.546 0,500 |
0,170 0.075 |
|
1—2 тыс.люкс |
ИУК, кинетин |
-5.5 |
20,3 |
0.583 |
0,118 |
|
|
ИУК. 6 БАП |
2,1 |
15,8 |
0,133 |
0,021 |
Большой интерес имеет селекционная интерпретация параметров фона для каллусогенеза и регенерации (таблица). Как правило, при оценке фона обычно исходят из критерия оптимальности процесса, т. е. наибольших значений признаков in vitro. На наш взгляд, такая установка не совсем обоснована и уместна лишь в тех случаях, если целью является не отбор, а культивирование генотипов in vitro. При клеточной селекции так же, как и при селекции в полевых условиях, важны типичность и дифференцирующая способность среды. При этом типичность можно понимать двояко: по отношению к условиям in vitro и по отношению к условиям in vivo. Во втором случае точкой отсчета должны быть количественные признаки, представляющие хозяйственную ценность. Первый вариант оценки типичности будет представлен в настоящем подразделе главы, второй — в следующем.
Анализируя параметры, мы будем понимать под "средой" сочетание условий культивирования (температура, освещенность) с составом питательных сред.
При оценке сред по эффективности процесса каллусогенеза очевидно преимущество оптимальной температуры, в особенности на среде с НУК. Последняя среда оптимальна для культивирования каллуса образцов томата. Однако она обладает наименьшей относительной дифференцирующей способностью, и по этому показателю выделяются среды при культивировании на пониженной температуре. Наиболее типичными были среды, содержащие НУК, а лучшими по предсказующей способности условия культивирования при пониженной температуре. Таким образом, "богатые" среды не имели преимущества перед "бедными" как фон для отбора.
Таблица Связь между признаками томата в культуре in vitro и in vivo.
|
№ п/п |
Признаки |
Номер признака |
|||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
||
|
1 |
Общий урожай |
1,0 |
|
|
|
|
|
|
|
|
|
|
2 |
Товарный урожай |
0.975 |
1,0 |
|
|
|
|
|
|
|
|
|
3 |
Ранний урожай |
0.106 |
0.078 |
1.0 |
|
|
|
|
|
|
|
|
4 |
Число плодов на кисти |
-0.195 |
-0.167 |
0,208 |
1.0 |
|
|
|
|
|
|
|
5 |
Завязываемость |
0,013 |
0,030 |
-0,364 |
0.140 |
1.0 |
|
|
|
|
|
|
6 |
Число кистей на главном стебле |
-0.256 |
-0,153 |
-0,625 |
0,330 |
0,377 |
1.0 |
|
|
|
|
|
7 |
Высота растения |
-0.073 |
0,024 |
-0,704 |
0,157 |
0,430 |
0,911 |
1.0 |
|
|
|
|
8 |
Длина пасынка |
-0.051 |
0,023 |
-0,678 |
0,276 |
0,482 |
0,874 |
0,932 |
1.0 |
|
|
|
9 |
Масса каллуса |
0,021 |
0.008 |
0,121 |
0,041 |
0,140 |
0,025 |
-0.089 |
-0.029 |
1.0 |
|
|
10 |
Регенерационная способность |
0,064 |
0,006 |
0,070 |
0,001 |
0,156 |
-0,156 |
-0.079 |
-0.101 |
0.189 |
1.0 |
Близкая картина наблюдалась при оценке по регенерационной способности. Лучшими были среды, содержащие ИУК и 6 БАП как при нормальной, так и при пониженной освещенности. Эти же среды имели самую низкую относительную дифференцирующую способность и слабую или среднюю типичность. По предсказуемой способности выделялись среды с ИУК и кинетином, в особенности при нормальной освещенности.
Анализ параметров фона по признакам томата в культуре in vitro позволяет сделать вывод, что понятия "оптимальная среда для культивирования" и "оптимальная среда для отбора" не совпадают. В последнем случае среда не обязательно отличается высокими значениями признаков генотипов в культуре in vitro. Успешный отбор может проводиться и на стрессовых средах при высокой дифференцирующей способности генотипа и типичности условий среды.
