Энергетическая эффективность сельскохозяйственного производства и факторы, ее определяющие
Методы оценки энергетической эффективности сортов и гибридов.
Сельское хозяйство является важным потребителем невосполнимой энергии, поэтому энергетический анализ эффективности агро- экосистем получает все большее распространение. Его достоинство в универсальности, что позволяет сопоставить эффективность любых элементов технологии (внесение удобрений, пестицидов, использование техники и т.д.). Для того чтобы давать корректную оценку тенденций изменения энергетических затрат в агроэкосистемах в процессе развития человеческого общества, необходимо выявить, как изменялась эффективность потребления энергии растениями в процессе эволюции и селекции.
Г. Одум, Э. Одум (1978) сформулировали закон максимизации энергии: в соперничестве с другими системами выживает та из них, которая наилучшим образом способствует поступлению энергии и использует максимальное ее количество наиболее эффективным способом.
Авторы пришли к выводу, что такая система:
- создает накопители высококачественной энергии;
- затрачивает часть накопленной энергии на обеспечение поступления новой энергии;
- активизирует круговорот различных веществ;
- создает механизмы регулирования, поддерживающие устойчивость системы;
- налаживает обмен с другими системами, необходимый для обеспечения потребности в энергии специальных видов.
И. И. Свентицкий (1982) обосновал принцип биоэнергетической целенаправленности структур и функций живых систем: "Каждый элемент живой природы в своем развитии и функционировании стремится к состоянию, обеспечивающему в существующих условиях внешней среды наиболее полное использование свободной энергии системой данного трофического уровня, в которую он входит". Автор считает, что выживаемость организмов и их приспособленность к условиям среды на протяжении эволюции определялись принципом биоэнергетической общей направленности.
По мнению И. И. Шмальгаузена (1972), в процессе эволюции организмы снижают энтропию, т.е. увеличивают свою упорядоченность в результате естественного отбора особей, наиболее успешно разрушающих внешнюю среду, т.е. повышающих ее энтропию. При этом в результате движущей формы отбора средством снижения энтропии является приспособление к максимальному использованию среды, а в результате стабилизирующей формы повышается экономичность этого использования путем увеличения устойчивости и упорядоченности наследственного кода, индивидуального развития и фенотипа.
Н. Д. Озернюк (1992) сформулировал принцип минимума энергии в онтогенезе, в соответствии с которым развивающийся организм, "выбирая" оптимальные условия существования, попадает в такие условия, в которых уровень его энергетического обмена минимальный (рисунок 16). Минимизация суммарных энерготрат организма во время онтогенеза включает минимизацию энерготрат на обеспечение процессов жизнедеятельности (ЕR) и процессов адаптации к неблагоприятным условиям среды (Еa): ЕR+Еa=min.
По мнению автора, минимизация энерготрат в онтогенезе является эволюционной адаптацией, отражающей общую стратегию взаимодействия организма и среды.
Однако в литературе мало сведений об изменениях энергетической эффективности растительных организмов в процессе селекции. Если учесть, что селекция от эволюции отличается прежде всего преобладанием движущей формы над стабилизирующей, можно предположить, что в процессе селекции, вероятно, повышается способность усваивать энергию, вносимую в агроценоз человеком (удобрения, поливы и т.д.), и увеличиваются затраты энергии, связанной с реакцией растений на биотические и абиотические стрессы. Иными словами, возрастает отзывчивость на регулируемые факторы среды и может падать устойчивость к нерегулируемым. Оба этих положения находят достаточно частое подтверждение в агрономической практике.
С каждым годом энергетические затраты в сельскохозяйственном производстве имеют тенденцию к возрастанию. Общеизвестен закон снижения энергетической эффективности природоиспользования: с ходом исторического времени при получении из природных систем полезной продукции.

Рисунок. Схема изменений энергетического обмена в онтогенезе.
По мнению автора, с начала века количество энергии, затрачиваемое на 1 единицу сельскохозяйственной продукции, в развитых странах мира возросло в 8 — 10 раз.
Повышение энергетических затрат сопровождается загрязнением окружающей среды пестицидами, нефтепродуктами и другими поллютантами. В связи с этим весьма важно выяснить структуру энергозатрат в растениеводстве, а также роль сорта как биологической системы по переработке энергии и возможности экономии энергозатрат в процессе совершенствования этой биологической системы селекционным путем.
В отличие от естественных экосистем, где единственным источником энергии является энергия солнца, агроценозы используют техногенную энергию (удобрения, топливо, пестициды и т.д.). По мнению Г. А. Булаткина, В. И. Ватолина (1979), доля антропогенной энергии от всей энергии, аккумулированной в урожае, составляет 5 — 10%. Наибольший удельный вес антропогенной энергии (17 — 19%) — в защищенном грунте как объекте наиболее интенсивного растениеводства (Свентицкий, 1982). По данным Л. М. Блянкмана, Н. И. Анисимовой (1990), затраты энергии в растениеводстве Беларуси в 1985 г. приходились главным образом на топливо (50,98%) и минеральные удобрения (41,39%). При этом часть энергии, накопленной в первичной продукции, теряется в результате действия болезней и вредителей (до 25%) и при перевозке и хранении (10 — 60%).
Для оценки энергетической эффективности сельскохозяйственного производства важно сопоставить энергию, накопленную в продукции и биомассе растения, с затратами антропогенной энергии. А. А. Жученко (1990) предлагает использовать для этих целей два показателя: коэффициент продуктивной (КПЭЭ) и общей (Коээ) энергетической эффективности:

По мнению автора, эгроэкосистемы позволяют получать на одну используемую техногенную килокалорию от 2 до 6 пищевых килокалорий. Как правило, повышение энергозатрат приводит к снижению энергетической эффективности производства. Так, в защищенном грунте Кээ составляет соответственно для огурцов и баклажанов 0,02 и 0,05 (Жученко, 1990).
Глубокий анализ физиологических и биохимических основ энергообмена растений предпринят Н. С. Балауром с сотр. (Балаур, Копыт, 1986, 1989, 1991; Балаур, Тетю, 1988; Балаур, Воронцов, Копыт, 1990). Н. С. Балаур выдвинул концепцию энергетического бюджета организма, сущность которой состоит в определении и регулировании генетически детерминированного количества энергии, необходимого для формирования продуктивности и устойчивости растений в конкретных условиях среды. При этом автор подчеркивает важность изучения энергообеспечения растений не только в оптимальных, но и в стрессовых условиях, что позволяет вычленить энергетическую "цену" адаптивной реакции генотипа на стрессовый фактор. По мнению Н. С. Балаура, повышение эффективности использования энергии растением — генетическая и селекционная проблема, предусматривающая создание форм растений с прямостоячими листьями, скороспелых и толерантных сортов, использующих воду и удобрения с большей эффективностью и способствующих связыванию большего количества солнечной энергии.
Предполагается на основе методов генетической инженерии создать формы растений с минимальными энергетическими затратами на биосинтез клеточного материала. При этом предусматривается сокращение множественных копий ДНК в хлоропластах и митохондриях, сведение к минимуму затрат на тем- новое дыхание, перераспределение части повторяющейся ДНК генома и т.д. Необходимо применять и традиционные методы селекции для отбора эволюционно сложившихся энергоэкономных генотипов, способных при меньших энергозатратах метаболической энергии формировать оптимальную продуктивность.
Для повышения энергетической эффективности возделывания культуры необходимо максимизировать выход энергии с продукцией и минимизировать энергетические затраты на ее производство.
При этом важно учитывать межвидовые различия растений по накоплению энергии в урожае. В. И. Кривченко и др. (1988) классифицировали растительный генофонд по содержанию энергии в единице хозяйственно ценной части продукции (таблица 101). Наименьшей энергетической эффективностью обладают овощные культуры, наибольшей — зерновые и зернобобовые. В. В. Коринец, Л. Н. Попова, В. И. Попов (1988) считают, что такой системно-энергетический подход можно применить в селекции растений. При этом следует учитывать разницу между сортами по энергонакоплению.
Таблица 101. Классификация растительного генофонда по содержанию энергии на 100 г продукции (по Кривченко и др., 1988).
|
Группа |
Энергетическая ценность. Дж |
Культура |
|
I |
До 50 |
Огурец, сельдерей и др. |
|
II |
50 — 100 |
Редис, томат, салат, шпинат, ревень, перец, лук (перо) и др. |
|
III |
100 — 200 |
Баклажан, перец красный сладкий, брюква, кабачки, капуста белокочанная, лук порей, лук репчатый, морковь, петрушка (корень), редька, репа, сельдерей (корень), укроп, фасоль, щавель, арбуз, дыня, тыква и др. |
|
IV |
200 — 300 |
Свекла, бобы и др. |
|
V |
300 — 700 |
Картофель, горошек, чеснок и др. |
|
VI |
700 - 1200 |
Рис и др. |
|
VII |
1200 — 1300 |
Овес, просо, гречиха, горох, фасоль и др. |
|
VIII |
1300 - 1400 |
Озимая пшеница, яровая пшеница, рожь, ячмень, кукуруза и др. |
|
IX |
Более 1400 |
Кукуруза (крахмалистая, восковидная), соя, арахис, кунжут, рапс (семена) и др. |
Сорт является не только основным средообразущим фактором в сельском хозяйстве, но и преобразователем солнечной и антропогенной энергии. От него зависят дозы удобрений и пестицидов, затраты топлива при обработке почвы, энергозатраты при переработке и хранении продуктов питания. Задача создания энергоэффективных сортов была поставлена в работах ряда авторов (Балинт, 1981; Hogenboom, 1983; Dambroth, Bassam, 1983 и др.). При этом важно давать оценку энергетической эффективности сортов. М. М. Севернев и др. (1991), М. М. Севернев (1992) предлагают для оценки энергетических затрат базового и нового сортов использовать следующие формулы:
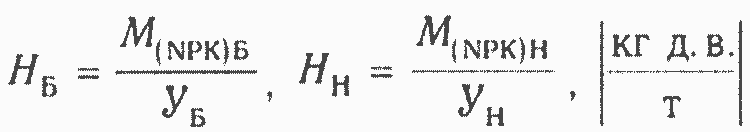
где УБ, Ун — урожай базового и нового сортов растений, т; МБ% Мн — затраты элементов питания по базовому и новому сорту, кг д.в.
Для оценки энергоемкости на единицу урожая используются энергетические эквиваленты каждого из видов удобрений α(N);α(P);α(K)
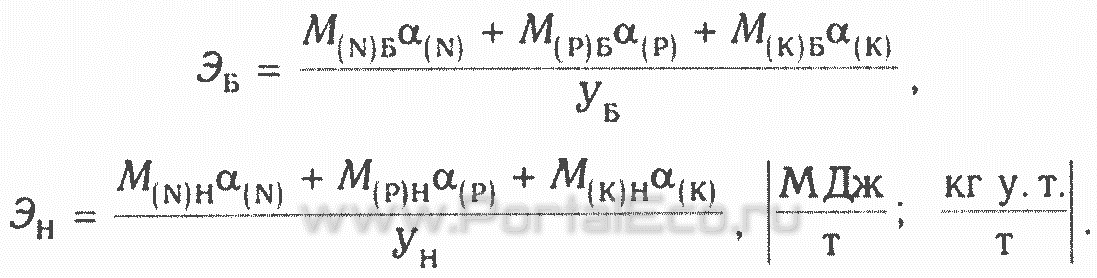
Общий коэффициент энергетической эффективности определяют по формуле Кэ=Эн/Эь. Аналогично можно рассчитать Кэ по другим затратам энергии (топливо, пестициды и др.).
В. И. Галкин (1981) предложил использовать для оценки отзывчивости сортов на регулируемые факторы среды два показателя: базовый и переходный коэффициенты эффективности использования фактора среды (#эи). Базовый Кэи — это количество граммов фитомассы растений, которое образовалось в расчете на 1 г NPK или 1 л поливной воды, переходный КЭУ[ — изменение величины признака при переходе от одного уровня фактора к другому, рассчитанное на единицу фактора. Автором проведен вегетационный опыт с 12 сортами пшеницы разной селекционной продвинутости из четырех экологических групп. Показано слабое влияние селекции интенсивных сортов на отзывчивость их к факторам среды. Выявлена различная сортовая реакция на факторы среды (удобрения, влажность почвы, густота стояния), которая зависела главным образом от экологического происхождения сорта. В селекции интенсивных сортов произошло повышение оптимального уровня минерального питания растений.
В селекции, как правило, оценивают сортообразцы, соблюдая принцип единственного различия. В этом случае коэффициент энергетической эффективности нового сорта в сравнении с базовым можно оценивать путем сравнения их урожаев. Такой подход не несет новой информации. На наш взгляд, правильнее давать оценку энергетической эффективности сортов на двух-трех агрофонах по приросту урожайности и/или биомассы на единицу дополнительных энергозатрат: Кээс=Эу/Эф
где Кээс — коэффициент энергетической эффективности использования фактора среды (удобрения, орошение и др.); Эу — дополнительная энергия от прибавки урожая; Эф — дополнительная энергия фактора среды.
При анализе ряда факторов среды можно определить суммарный Кээс: Кээс=ΣЭу/ΣЭф.
Таким образом, можно выявить сортовую отзывчивость на любой фактор среды.
Известно, что такая отзывчивость находится под генетическим контролем и может быть улучшена в процессе селекции. Создание энергетически эффективных сортов требует изучения исходного материала по отзывчивости на регулируемые факторы среды, генетического анализа такой отзывчивости, выбора модели сорта для конкретного уровня энергозатрат (дозы удобрений, орошение и т.д.), подбора фонов для выделения нужных генотипов в селекционном процессе.
Для этого необходимы глубокие исследования генетики и физиологии отзывчивости растений на регулируемые условия среды как на уровне изучения признаков, так и путем анализа энергетической цены отзывчивости в оптимальных и стрессовых условиях роста растения.
